Systematics
Author: Theodore R. Simons (TS), USGS NC Cooperative Fish and Wildlife Research Unit , Dept. Biology, NC State University, Raleigh, NC.
Geographic Variation
White is absent or nearly absent from the primaries of Pacific birds, but present at the base of several inner primaries of Atlantic birds. Birds in w. North America, on the Galapagos, and in se. South America have the dorsum dark and demarcation between dark and white on lower breast mottled, whereas birds in w. and n. South America and in e. North America have the dorsum browner and the demarcation on the lower breast crisp. The largest birds, on average, are in w. North America and the smallest in w. South America, whereas birds on the Galapagos have disproportionately “larger” legs and toes (Hayman et al. 1986) and birds in se. South America average the shortest tarsi. Some geographic variation may be the result of persistent hybridization with H. bachmani Audubon, 1838—the Black Oystercatcher of w. North America (see Jehl 1985)—and H. ater Vieillot and Oudart, 1825—the Blackish Oystercatcher of w. and s. South America (Hayman et al. 1986, Hockey 1996a).
Subspecies
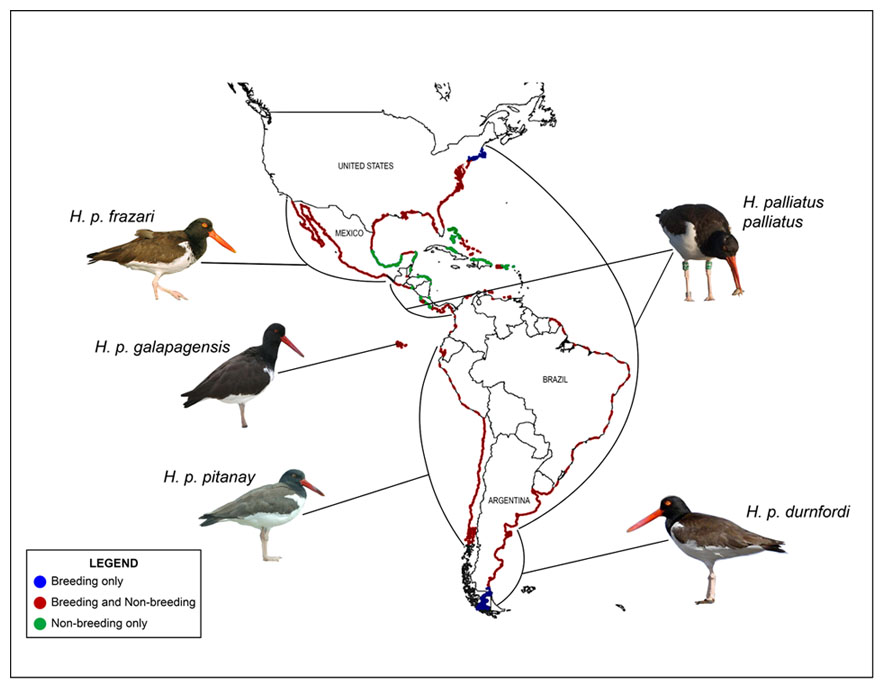 Five subspecies (Ridgway 1919, Jehl 1985; see Distribution), although that total is disputable. There may be as few as two subspecies (Hockey 1996a), and Hayman et al. (1986) noted, on the basis of Allan J. Baker’s input, that there may be “no valid subspecies . . . that the observed pattern has resulted from clinal variation and hybridisation with black oystercatchers,” in this case meaning with H. bachmani and H. ater. A comprehensive and quantitative analysis of geographic variation is needed.
Five subspecies (Ridgway 1919, Jehl 1985; see Distribution), although that total is disputable. There may be as few as two subspecies (Hockey 1996a), and Hayman et al. (1986) noted, on the basis of Allan J. Baker’s input, that there may be “no valid subspecies . . . that the observed pattern has resulted from clinal variation and hybridisation with black oystercatchers,” in this case meaning with H. bachmani and H. ater. A comprehensive and quantitative analysis of geographic variation is needed.
H. p. palliatus Temminck, 1820. Includes H. hypoleuca Pallas, 1811; H. arcticus Jardine, 1832; H. brasiliensis Wied, 1821; H. p. prattii Maynard, 1899. Resident along Atlantic coast of North America from New Jersey (breeding north to Massachusetts) south to central Florida, along nearly the whole coast of the Gulf of Mexico (where apparently non-breeding only in the southern fringe), and around the s. Caribbean Sea south and east along the coast of n. South America, including the Netherlands Antilles; also in The Bahamas and locally in the Greater Antilles and leeward Lesser Antilles and on the Pacific coast of s. Mexico and Central America [type locality = Venezuela]. Dorsum dark brown; uppertail coverts clean white; base of inner 2–4 primaries white; crisp line between dark hood and white ventrum.
H. p. frazari Brewster, 1888. Resident in w. Mexico from n. Baja California (rarely to s. California) and along w. Mexico south to the Isthmus of Tehuantepec [type locality = Isla Carmen, Gulf of California]. Resembles H. p. palliatus but dorsum darker (blacker), uppertail coverts mottled (especially laterally, rendering the white rump patch much smaller), inner primaries generally lack white, and demarcation between dark hood and white belly mottled, with dark sometimes extending into the center of the upper belly (Jehl 1985, Hamilton et al. 2007); averages larger (e.g., mean bill length = 75 mm). The blackish dorsum, marbled uppertail coverts, and blurred demarcation line on the breast may be the result of genetic introgression from hybridization with H. bachmani (Brancroft 1927, Jehl 1985), whose range overlaps that of H. palliatus on the Pacific coast of n. Baja California (and sometimes s. California, especially the Channel Is.; see Hamilton et al. 2007).
H. p. galapagensis Ridgway, 1886. Resident on the Galapagos Is. [type locality = I. San Cristóbal (a.k.a. Chatham I.)]. Like H. p. frazari but uppertail coverts clean white; dorsum averages darker, and averages longer toes and shorter wings and tail (see Ridgway 1919).
H. p. pitanay Murphy, 1925. Resident along Pacific coast of South America from Ecuador to Chile [type locality = Pisco Bay, Peru]. Resembles H. p. palliatus but white absent from primaries; averages smaller.
H. p. durnfordi Sharpe, 1896. Replaces H. brasiliensis Wied, 1821, a nomen nudum. Resident along the Atlantic coast of South America from s. Brazil south to central Argentina [type locality = mouth of Chubút R., Argentina]. Resembles H. p. frazari but bill (mean = 65 mm) and legs short and white present at base of primaries.
Related Species
On the basis of morphology and various genetic tools, family Haematopodidae, the oystercatchers, is phylogenetically nearest family Recurvirostridae, the stilts and avocets (Thomas et al. 2004, van Tuinen et al. 2004). Haematopodidae is itself distinct, but the various species in the family are remarkably uniform in structure and basic plumage pattern (see Distinguishing Characteristics), whether wholly black or pied—although some species are dimorphic. As a result, species-level taxonomy remains unresolved and even controversial, with as few as four and as many as eleven species recognized (Murphy 1925, 1936, Peters 1934, Heppleston 1973a, Hayman et al. 1986, Hockey 1996a, Hebert et al. 2004, A. J. Baker pers. comm.). In this vein, H. palliatus has been treated as conspecific with H. bachmani and with H. ostralegus, the Eurasian Oystercatcher (e.g., Peters 1934, Murphy 1936, Heppleston 1973a). Ongoing hybridization between some species clouds the picture, including between H. palliatus and both H. bachmani in s. California and nw. Baja California and H. ater in s. South America (Jehl 1985, Hockey 1996a).